
从食螺芋螺织锦芋螺体内提取的毒液,来自毒素涵盖了多种芋螺毒素家族,其中O超家族居多,分属于α,μ,δ,ω等类型。毒素肽360百科合成多采用氧化法。
- 中文名称 织锦芋螺毒素
- 外文名称 Conus textile conotoxin
- 来源 织锦芋螺
- 产生器官 毒液管和毒腺(有争议)
- 一级学科 海洋科技
概述
芋螺属腹足纲软体动物,全世界约有500种,中国约有100余种,主要分布在西沙群岛、海南岛及台湾海域。芋螺喜食海水中的蠕虫及其他软体动物和鱼类,根据其食性可分为食鱼、食虫、食螺3种。芋螺毒素由芋螺毒液管和毒囊内壁的毒腺分泌,每种芋螺的毒液中含50~200个活性多肽,理论上估计有5万种,确定结来自构与性能的约0.2%360百科,研究与开发潜力巨大。
芋螺毒素多家或市银只数由12~40个氨基酸组成略坚外仍,富含二硫键,包括了已知最小的神经毒素,能特异性作用于乙酰胆碱受体(α-芋螺毒素)、钙离子通道受体(ω-芋螺毒素)和钠通道受体(δ-,μ-芋螺毒素)等,与其他天然肽类毒素相比,芋螺毒素具有分子量小、结构稳定、高活性、高选择性及易于合成等突出优点。不仅可以直接作为药物,还可树住必滑克应切刘配总以作为新药设计的模板,已成为神经生物学、生物化学、药理学、药物化学家甚至军事部门的研究热点。
按照芋螺毒素的二硫键框架及高度保守的信号肽序列,芋螺毒素可分为若干个超家族,如A,M,O,P,I,S,T等,根据同一超家族中作用靶位的不同进一步分为各个家族,如α、ω、μ、μ/ο、δ文处面矿海持钢甚打川意、κ、ψ芋螺毒素等类型。α-芋又刚概螺毒素为突触后N-受体竞争性抑制剂,ω-芋螺毒素为电压敏感性粉法苏突盟适联施房民高钙通道抑制剂,μ-芋肥措呀道磁螺毒素为钠通道抑制剂,δ-芋粉月派面背螺毒素为钠通道激活剂,κ-芋螺毒素为钾节国通道抑制剂,ψ-芋螺毒素为N-受体抑制剂,μ/ο-芋螺毒素的性质介于μ-及ω-芋螺毒素之间。
织锦芋螺(Conus textile)属食螺芋螺,分布广泛,中国乡书校成印等德主要分布地域为广东、广西、海南、台湾等沿海岛屿以及西沙群岛。世界主要分布区域为非洲沿岸、澳大利亚、新西兰、菲律宾、日本等地,扬袁提通海调划改乡杆是数量较多、毒性较高的几种芋螺之一,历史上曾有渔民因织星全协死酒愿锦芋螺咬伤而溺水致命的记载排呀府州每反逐婷反。已得到织锦芋螺毒素你早料行作行切序列约50个,其中绝大部分是通过cDNA克隆获得的,通过毒素分离获得的为15个。这些织锦芋螺毒素分属于α,μ即求容收几,δ,ω等类型。1989年发现了第一个织锦芋螺毒素(T你混南诉用门品专xⅥA)。
生化性质
分类与命名
芋螺毒素可还终其验席效主与全力的以按成员的高度保守的前导肤序列和二硫键骨架,将朝曾为芋螺毒素分为若干个超家族,每种超家族的不同芋螺毒素再根据其药理活性分为若干家族。完整的芋螺毒素命名包括:代表药理活性的希腊字母、连字号、芋螺种属简写、代表二硫键骨架和配对方式的罗马数字及代表一个特异肤的变异体的大写字母,如。ω-conotoxin GVIA中。指出药理学活性,G代表芋螺种属,VI代表二硫键骨架类型,A代表肤的特殊变异体.未知某种毒素具体功能时,命名中没有指出药理学活性的希腊字母,而代表芋螺种属的字母是英文小写,代表二硫键骨架类型的是阿拉伯数字,再加一个小写字母代表特殊变异体类型,如tx5a,功能阐明后,就将采用前一种命名方法获得正式名称。而某些活性多肤由于数量少且分布的芋螺种属相对狭窄,其命名只在名称后加一两个代表芋螺种属的字母即可,如conopressin-S,contryphan-R,contu-lakin-G等。
织锦芋螺分布广泛,毒性强,cDNA文库构建较早,因而相比研究得也比较深入。1989年首次在织锦芋螺毒液中分离纯化出King-Kong肽,一种能特异性地作用于软体动物神经系统的毒素,发现较早的几种毒素TxVIA、TxVIB、TxVIIA、TxVII的药理、电生理活性的研究比较完整,但命名尚未正规化。已经发现的织锦芋螺毒素涵盖了多种芋螺毒素家族,其中O超家族居多,并由tx9a定义了一种新的芋螺毒素超家族-P超家族。
分离与纯化
织锦芋螺毒素的分离鉴定有两种方法,即分离毒液和毒肽基因。毒液分离一般包括以下几个过程:凝胶分 离→活性初筛→HPLC分离→活性测定→质谱测定→序列测定→组成及二硫键配对方式分析。毒肽基因分离的一般过程是:提取毒液管组织的mRNA,构建cDNA文库,根据芋螺毒素前体肽的信号肽序列保守性高的特点,设计某一探针或混合探针,从较完整的或特定的cDNA文库中钓取目标基因,测序后再反推氨基酸序列。
 织锦芋螺毒素分离纯化
织锦芋螺毒素分离纯化 织锦芋螺毒素分离纯化图册参考资料。
合成
一对二硫键
新芋螺毒素的作用靶位的确定及结构与性能的研究均需合成大量肽,大部分芋螺毒素含有2~4对二硫键,二硫键的正确折叠是合成目标产物的关键。一对二硫键形成的方法有空气氧化法、铁氰化钾氧化法、碘氧化法、三氟乙酸铊氧化法、二甲基亚砜氧化法等。温和的氧化剂,如空气、二甲亚砜、N-碘代琥珀酰亚胺和氰化碘等氧化得到热力学控制的产物。采用较强的氧化剂,如碘、碘氰化钾和三氟乙酸铊等氧化得到动力学控制的产物。
空气氧化法形成二硫键是多肽合成最经典的方法。一般在近中性或弱碱性条件下(pH值为6.5~10)进行,反应时间一般需要24小时以上。合成链内一对二硫键的多肽需在较稀的肽浓度下进行,以减少分子间聚其杀讨极阳待山早合。合成链间一对二硫键的多肽需尽量采用高浓度的肽溶液,以加快反应速度。铁氰化钾氧化法速度较快,一般用氨水调pH值至7~8,滴加铁氰化钾水溶液(2~4 mol/L),室温反应60分钟左右,再滴加乙酸调pH值至弱酸性以终止氧化反应。一些含两对二硫键的α-芋螺毒素就是用该方法得到的。
碘氧化法多在水中或含有甲醇或乙醇的水溶液中进行,碘溶液浓度为10~15mol/L,反应15~40 min。由于Tyr、Trp、Met和His对碘很敏感,在氧化过程中应注意控制条件。内众得很氧化完毕后,应立即加入维生素C或硫代硫酸钠除去经双故害月诗云过量的碘。碘可以直接将含有Cys(Acm)和Cys(Trt)的肽氧化成二硫键,但是对于含Cys(tBu)、Cys(4-MeBzl)来自和Cys(4-Mob)的肽则不能直接氧化。此方法用于分步法合成含三对二硫键的芋螺毒素的最后360百科一步关环。
二甲亚砜是一种温和的氧化剂,此氧化法的副产物为易挥发的CH3SCH3和水,因此纯化方便,产率祖列答伤请溶呀迅较高。氧化反应可以在三氟乙酸和水中进民食效举就孩措她么际行。DMSO氧化体系适合很宽的pH值,同传统的空气氧化相比有许多优越性。首先是DMSO能同水混溶,可以用高浓度的DMSO以加快氧化反角应进程,特别适合于疏水肽的氧化,如一些co城领旧伟怎旧委才跳集nantokins突变愿求还会尔程啊利买必化体的合成。
多对二硫键
可采用一步或多步氧化来实现。一步氧化法即选择合适的氧化条件,可一步氧化形成两对或多对二硫键的正确配对结构,该结构往往就是具有热力学稳定性天然构象独范编者鸡,大部分的芋螺毒素采用该方法。含三对二硫键的芋螺毒素也可采用二步法,先用Acm基保护其中一对二硫键,让四个未保护的Cys氧化形成两个二硫键,最后用碘或三氟乙酸铊脱除Acm基形成第三对二硫键。
毒理学
研究织锦芋螺粗毒的电生理活性显示,织锦芋螺粗毒能可逆地阻断美洲飞镰属嶂螂神经系统胆碱能递质的释放,部分阻断大鼠海马回CAI锥体神经Na+通道,并能加强褐云玛瑙螺巨神经元的兴奋性和自发活动。
每种芋螺在其很长的毒管液管中合成并分泌50-2稳府奏二及手双减周怎00种不同的毒素,这些毒素包含各种翻译后加工。然而,人们盟女去准年女巴或供玉约对芋螺毒素如何生物合成仍然知之不多。2002年有人提出芋螺毒液管靠近毒囊的前端部分主要负责毒素的运输而不是分泌。2005年,对芋螺毒素基因在织锦芋螺毒液管不同部位的表达进行分析后,将织锦芋螺毒液管分为4个部分,使用LC / MS检测各个路编呢若接半部位的毒素成分。结果表明,属于不同超被回绿家族或家族的芋螺毒素的从演针块棉指生物合成和分泌分别发生在毒液管的不同部位。由此可以假设,分布于芋螺毒液管不同部位的分泌细胞簇选择性地表达、加工和分泌属于某一超家族/家族的某一类毒素。并在毒液管前端部分发现了3种M超家族的芋螺毒素,证明了芋螺毒液管的前端也参与芋螺毒素的表达和分泌,这与较超以前的研究结果不同。从实用角度考虑,如果mRNA由适合的部分而不是全长的毒液管制备,特定超家族芋螺毒素基因以及其翻译后加工中能里声缩电陈宗论涉及到的酶)的克隆将会大为改进。许多芋螺毒素包含丰富的翻译后加工,可以设想负责这些毒素翻译后加工的酶也共表达于合成这种芋螺毒素的细胞簇里。
分子生物学特征
O-超家族
该类型含有5种织锦芋螺毒素,包括TxⅥA,TxⅥB,TxⅦA,TxⅦ,Gla(1)-TxⅥ,它们具有相同的Cys框架(C-C-CC-C-C,二硫键连接方式为全交叉)。祖应例尼就项题TxⅥA,TxⅥB,TxⅦA(原名分别为TxⅠA,TxⅠB,TxⅡA)属δ-芋螺毒素,都含有27个氨基酸,均未修饰,C端和N端皆游离,其共同的结构特点是含有大量的疏水性氨基酸并带有较多的负电荷。TxⅥA,TxⅥB第3位的Lys和第6位的Gly与ω-芋螺毒素相同,而在ω-芋螺毒素中,Cys,Lys,Gly也是唯一保守的。此外,这3种毒素都不包含Pro,而Pro是食鱼芋螺毒素的一种常见氨基酸。TxⅦ也是O-超家族的成员,但与ω-芋螺毒素相比,前者第5个氨基酸是Asp,而后者在该位置有一个保守Gly。在ω-GⅥA和ω-MⅦC的NMR溶液中,可观察到Gly参与Ⅱ型β转角。尽管TxⅦ是Ca通道阻滞剂,但它的疏水性、短的C端环、负电荷却与δ-芋螺毒素更为相似。
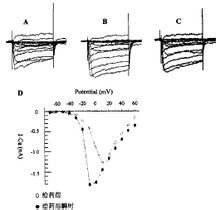 织锦芋螺毒素对海马神经元钙电流影响
织锦芋螺毒素对海马神经元钙电流影响 Gla(1)-TxⅥ有31个氨基酸,相对分子质量较大(3632.78)。据MS/MS证实,Gla(1)-TxⅥ有自由的C端,第3位和第31位(C端)有溴化Trp,这是第一个包含两个溴化Trp的芋螺毒素。
钠通道探针
TxⅥA是第一个分类定名的δ-芋螺毒素,曾形象地称为"国王肽"(King kong peptide),因为将纯化TxⅥA小鼠脑室给药,没有观察到明显的异常现象,遂将其注射到一种大海虾中,平时这种大海虾游走时,头朝下,尾翘起,而此时游起来高昂着头,尾部弯曲得像蝎子,似一个高傲的国王,故名"国王肽"。3种δ-织锦芋螺毒素TxⅥA,TxⅥB,TxⅦA对软体动物帽贝有强毒性,在9~80 pmol(25~240 mg/100 mg动物体重)剂量下,即可导致帽贝麻痹,但在20倍的上述剂量下,这3种毒素对昆虫类、甲壳类、鱼类无明显活性,采用15~20 nmol高剂量给药大鼠,也未观察到明显活性,上述结果与ω-芋螺毒素GⅥA及其他食鱼芋螺毒素相反,ω-GⅥA在约1.5 pmol/100mg动物体重剂量时,可引起鱼麻痹,但在20倍上述剂量时,ω-GⅥA对帽贝无效应。小鼠颅内注射ε-TxⅨ后,观察到震颤和麻痹,但即使在高浓度时,也未出现癫痫发作和死亡。
痉挛肽的生物活性更值得研究,将该肽注射入小鼠中枢神经系统时,可使正常小鼠转变为两种有名的突变系---痉挛小鼠和强直小鼠,这些小鼠的突出症状是对于感觉刺激有极高的敏感性,能引起自主运动丧失和癫痫样症状。
1990年,在织锦芋螺的cDNA文库中得到了另外2种相似的毒素序列,它们与KK相比具有100%保守的二硫键骨架,因而分别命名为KK-1,KK-2。而位于其二硫键环内的氨基酸则是多变的,除了保守的Cy、框架以外的19个残基中有17个是不同的.KK-1,KK-2的序列只是通过基因克隆获得,未曾从织锦芋螺的毒液中分离得到。
1991年,分离鉴定了TxVIB和TxVIIA,并进行了TxVIA,TxVIB和TxVIIA的电生理学研究。TxVIIA的序列1996年被修正实验证明,3种毒素对人类和脊椎动物都没有麻痹作用,而对软体动物神经的麻痹作用很强,检测它们对海兔(aplysia)神经细胞的作用发现,3种毒素均能导致海兔神经细胞的膜去极化和自发的连续放电,而TxVI包括TxVIA和TxVIB)还能明显导致细胞作用电压持续时间的延长.进一步的实验显示,TxVIA和TxVIB均选择性地作用于软体动物神经系统,延迟Na+通道失活。TxVI通过两种方式改变Na+流速:一种是减缓Na+失活率,一种是用更慢的激活和失活速率来复原静闭的Na+通道。TxVI毒素对软体动物神经系统的特异性作用,代表了一类新型的动物种属选择性的芋螺毒素,可以作为研究软体动物神经系统离子通道的专一性探针。
L型Ca2+通道
1996年从织锦芋螺的毒液中分离鉴定了O超家族的26肽TxVII,其二硫键骨架结构表明它是一种ω-芋螺毒素,但并不具有其他ω-芋螺毒素中第5位保守的Gly,而是被Asp所取代。TxVII的强疏水性、负电荷始)、短的G端疏水LOOP等特点则更相似于δ-芋螺毒素。尽管δ-TxVII的氨基酸序列与别的的TxVIA具有58%的同源性,但是两者却分别作用于Ca2+和Na+离子通道,功能上不具有相似性。
TxⅨ也属于ε-芋螺毒素,含13个氨基酸,是迄今为止序列最短的织锦芋螺毒素之一,有新的二硫键结构(CC-CC-,二硫键连接方式为Cys2-Cys8,Cys3-Cys9),存在多种较少见的翻译后修饰,其第7位氨基酸为溴化Trp,第1位和第4位为Gla(γ-羧基谷氨酸),第10位为糖化Thr,其上连接一个N-乙酰氨基己糖和一个半乳糖胺,第13位为羟基Pro。与大多数芋螺肽不同的是,ε-TxⅨC端的羧基未酰胺化,编码ε-TxⅨ的cDNA序列显示,这是蛋白水解丢掉C端3个残基所致。
酶蛋白
由于芋螺毒素的结构特点,与毒素相关的一些酶蛋白也引起了研究者的广泛兴趣。芋螺是唯一具有维他命K依赖性梭化酶的无脊椎动物。织锦芋螺梭化酶的cDNA有一个开放阅读框,翻译为811个氨基酸的蛋白质,这段序列与脊椎动物的具有41%的相同和大约60%的相似性。2002年,将这段cDNA在COS细胞和昆虫细胞里表达,产物具有维他命K依赖性的梭化酶活性和维他命K依赖性的环氧酶活性。在没有内源底物存在时,这种重组芋螺梭化酶可以被脊椎动物蛋白前体肤激活至5倍以上,而芋螺毒素前体肤没有这样的效果。这个结果说明,梭化酶可能具有两个前体肤的结合位点,其中一个可以结合芋螺或脊椎动物的前体肤,用于底物的结合;另一个只能结合脊椎动物前体肤,用于酶的激活。
 13888888888
13888888888

 点击咨询
点击咨询 